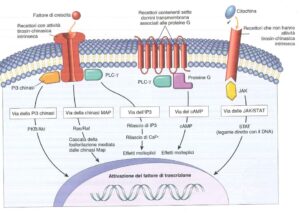
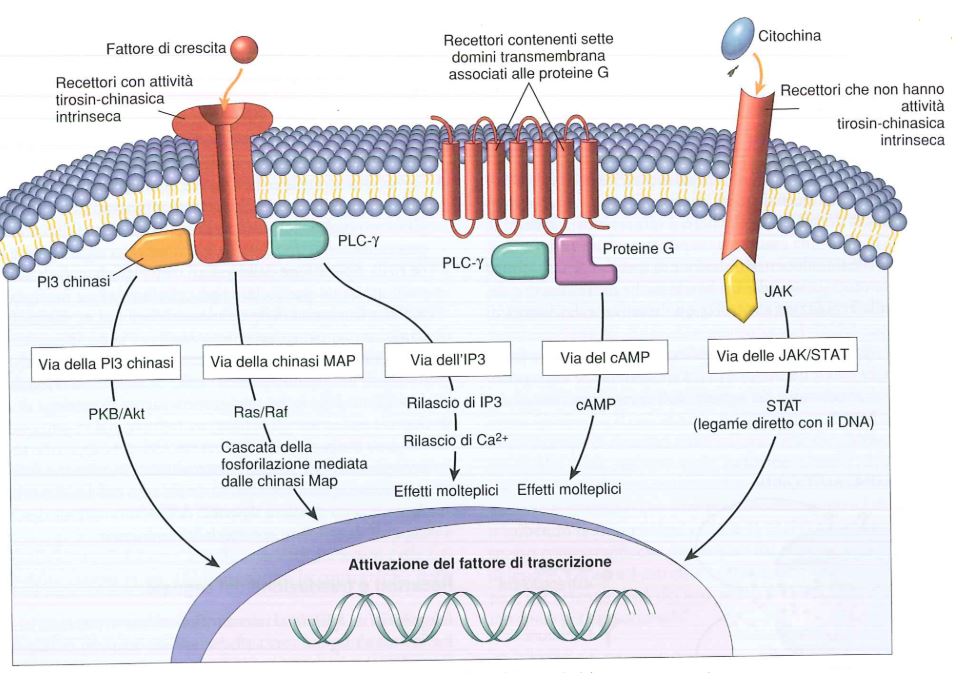
I messaggi chimici inviati dalla cellule, tra cui ormoni e fattori di crescita, hanno la capacità di interagire con dei recettori presenti sulla superficie della cellula stessa. Possiamo avere:
- recettori ad attività tirosin-chinasica intrinseca;
- recettori associati a proteine G;
- recettori che non sono dotati di attività tirosin-chinasica intrinseca.
Il recettore per il fattore di crescita deve interagire con un ligando che è il fattore di crescita stesso, e quando il fattore di crescita interagisce con il recettore, si deve scatenare una risposta intracellulare che si traduce in una determinata espressione genica, mentre la risposta finale deve essere la crescita della cellula.
Fonte: Le basi patologiche delle malattie

Recettori ad attività tirosin-chinasica intrinseca
I recettori ad attività tirosin-chinasica intrinseca sono caratterizzati da una porzione extracellulare, una transmembranaria e una intracellulare; quest’ultima è costituita da due porzioni che possono autofosforilarsi a livello della tirosina (da cui il nome). È un recettore monomerico che, nel momento in cui vi è interazione con il fattore di crescita, dimerizza, così è in grado di scatenare una auto-fosforilazione di tirosine che sono presenti nella faccia citoplasmatica del recettore stesso.
La fosforilazione della faccia interna è importante perché può far sì che il recettore medi una risposta intracellulare fungendo da punto d’ancoraggio per altre proteine adattatrici che si incaricano di trasferire il messaggio (attraverso ulteriori proteine di segnalazione) all’interno del nucleo.
Se ci fossero alterazioni a livello di questi recettori, si andrebbe incontro a patologie abbastanza serie, infatti queste alterazioni sono presenti nel cancro, diabete di tipo II, aterosclerosi e diverse altre patologie.
La risposta può essere mediata in vari modi.

Via delle proteine adattatrici
Questa via è seguita per esempio da: EGF (Fattore di Crescita Epidermico); TGF-α (Fattore di Crescita Trasformante); HGF (Fattore di Crescita degli Epatociti); PDGF (Fattore di Crescita per le Piastrine); VEGF (Fattore di Crescita VasoEndoteliale); FGF (Fattore di crescita dei Fibroblasti); c-KIT (coinvolto nella proliferazione ad esempio dei linfociti T); insulina.
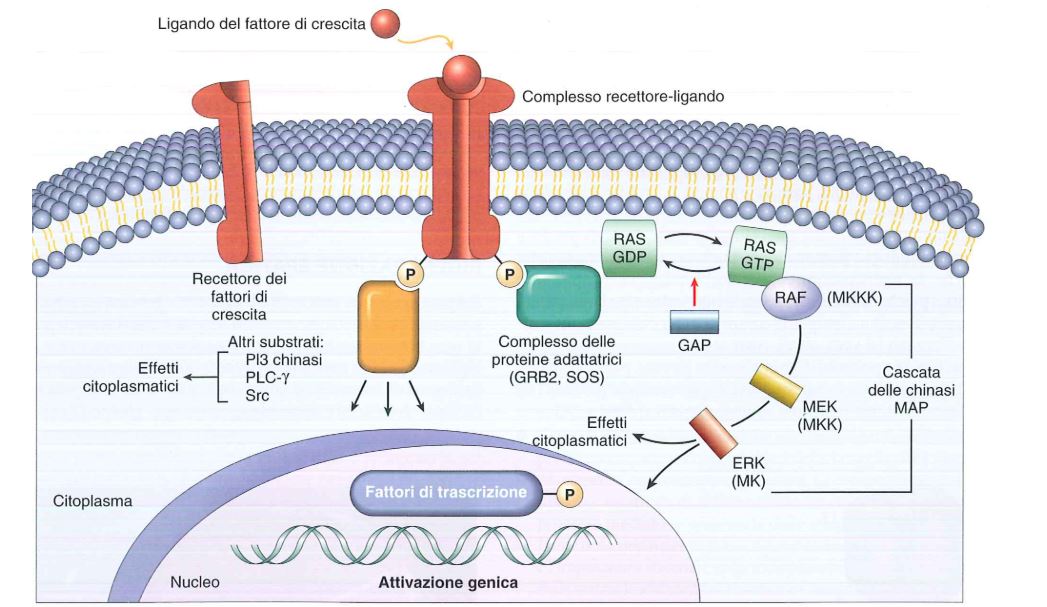
Le tirosine fosforilate possono legarsi a delle proteine adattatrici, tra cui le proteine SOS (Son of Sevenless) e GRB2 (Growth factor receptor-bound protein 2), che esaltano l’attivazione di altre proteine come le proteine Ras (Rat sarcoma). La proteina adattatrice riconosce il versante fosforilato e a sua volta interagisce con la proteina Ras che nella forma inattiva è legata al GDP (guanosina difosfato), nella forma attiva si lega alla GTP (guanosina trifosfato). La ras-GTP (un nucleotide guanilinico trifosfato) è importante perché è la forma attiva che può scatenare una risposta intracellulare che in ultima analisi porta all’espressione di determinati geni che servono alla crescita cellulare. Questo perché ras-GTP attiva a sua volta una cascata di MAP chinasi intracellulari che sono RAF (Rapidly Accelerated Fibrosarcoma), MEK e ERK, che a loro volta fosforilano due fattori trascrizionali: JUN e FOS, che nel nucleo dicono al materiale genomico di esprimere geni che servono per la crescita cellulare.
In questa via, GAP (GTPase Activating Protein) è un oncosoppressore in quanto dice a ras di staccarsi dal GTP e quindi di inattivarsi e inibire la crescita cellulare.
Quindi una risposta extracellulare di un fattore di crescita si è tradotto in un messaggio intracellulare di avanzamento della proliferazione.
Il recettore ad attività tirosin-chinasica intrinseca è importante perché può scatenare una via di messaggi intracellulari attraverso anche altri meccanismi come la via della PI-3-Chinasi.
Fonte: Le basi patologiche delle malattie.
Caricamento….
Via della PI-3-Chinasi (Akt/PKB)
Un altro meccanismo attraverso cui il recettore ad attività tirosin-chinasica intrinseca può scatenare una risposta intracellulare è la via della PI-3-chinasi.

Dopo la fosforilazione dei residui tirosinici del recettore ad attività tirosin-chinasica intrinseca in seguito al legame con il fattore di crescita, la PI-3-chinasi (fosfatidilinositolo-3-chinasi) si ancora alla porzione fosforilata del recettore. Questa, praticamente, attacca un fosfato a livello del fosfatidilinositolo difosfato PI(4,5)P2 (fosfolipide di membrana) nella membrana cellulare formando fosfatidilinositolo trifosfato, cioè PI(3,4,5)P3.
Questo meccanismo serve perché deve ancorarsi la PDK1 (Phosphoinositide-Dependent Kinase-1), una chinasi, a questi fosfati. Anche Akt o protein-chinasi B (PKB) si attacca ad un’altra molecola di PIP3. Precisamente abbiamo un fosfatidilinositolo trifosfato a cui si attacca PDK1 ed un altro vicino a cui si lega Akt. Essendo vicine sulla membrana, PDK1 catalizza la fosforilazione di Akt, che si attiva e può andare nel nucleo.
La PKB attivata media risposte di proliferazione cellulare perché favorisce l’attivazione di BAD fosforilandola (BAD appartiene alla famiglia delle Bcl-2 ed è un’antagonista della morte cellulare). BAD in forma inattiva è legata ad una proteina inibitrice. Se interviene la PKB attiva, BAD si stacca dall’inibitore e si lega alla proteina 14-3-3, mentre il dominio inibitorio precedentemente legato a BAD è in grado di inibire la risposta apoptotica. Quindi tutto questo segnale si traduce in una inibizione della risposta apoptotica e quindi nel proseguimento della crescita cellulare.
Caricamento…
Recettori associati a proteine G

I recettori associati a proteine G rappresentano la più vasta famiglia di recettori membranali (più di 1.500). Posseggono due domini elicoidali che attraversano 7 volte la membrana cellulare. Le molecole che possono utilizzare questo tipo di recettori sono: istamina, adrenalina, noradrenalina, rodopsina, vasopressina, serotonina, calcitonina, glucagone, paratormone, corticotropina, e vari farmaci. Esistono diverse vie.
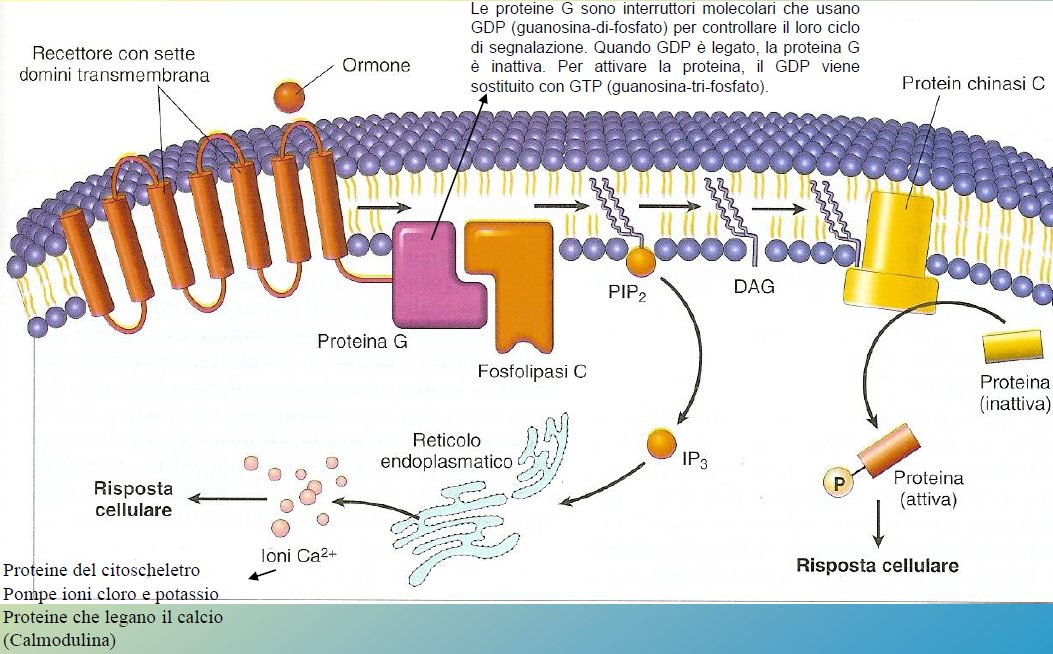
Via della PLCγ
Nel momento in cui abbiamo l’interazione con un ormone o con il fattore di crescita, si ha una attivazione di proteine G, che si trovano vicino al recettore a sette domini transmembrana. La proteina G è costituita da 3 subunità: α, β, γ. A seguito dell’attivazione la subunità α si lega al GTP, le subunità β e γ si distaccano e andranno ad attivare altre proteine (come per esempio la Fosfolipasi C, PLC).
La fosfolipasi C scinde i fosfolipidi. La fosfolipasi C-b (più precisamente), è in grado di scindere il PIP2 e formare l’inositolo trifosfato (IP3) e il diacil-glicerolo (DAG), che sono due importanti secondi messaggeri intracellulari. IP3 dice allo ione calcio (Ca2+) di uscire dal reticolo endoplasmico dove è accumulato (il calcio è sequestrato perché è pericolosissimo nel citoplasma, infatti fisiologicamente la concentrazione è 0,1 μM) ed attivare delle proteine calcio-dipendenti, come la calmodulina, cosicché si abbia la crescita cellulare. IP3 può far aumentare la concentrazione di calcio citosolica fino a 1 μM. Il calcio intracellulare è un importante segnalatore biochimico perché può agire può agire sulle proteine del citoscheletro, sulla calmodulina, sulle pompe cloro e potassio o può mediare una serie di segnali.
Invece, il diacil-glicerolo è in grado di attivare una protein-chinasi C, che a sua volta attiva altre vie di segnalazione il cui scopo ultimo è la proliferazione cellulare (via della protein-chinasi C).
Questa via è utilizzata anche dai recettori ad attività tirosin-chinasica intrinseca.
Via del cAMP
Un altro meccanismo attraverso il quale i recettori a 7 domini transmembrana possono dare una risposta di proliferazione cellulare è rappresentato da un altro sistema in cui, l’interazione del ligando con il recettore, porta alla attivazione della proteina G che attiva l’adenilato-ciclasi.
L’adenilato-ciclasi è in grado di trasformare l’ATP in cAMP (AMP-ciclico), il quale attiva la protein-chinasi-A (PKA), che quando è inattiva è legata a subunità inibitorie. Nel momento in cui si lega al cAMP, che stacca le subunità inibitorie, diventa attiva e va nel nucleo, dove si lega alla proteina CREB attivandola tramite fosforilazione. Questa proteina funge da fattore per la trascrizione di determinati geni che servono per la proliferazione cellulare.
Il cAMP regola anche canali ionici importanti nella vista e nell’olfatto.
Recettori non dotati di attività tirosin-chinasica intrinseca
Questi recettori si chiamano in questo modo poiché l’attività tirosin-chinasica è data da proteine distinte dal recettore stesso (Tyk2 e Jak1) che si appoggeranno ad esso. Sono importanti se consideriamo gli interferoni (interferone-α) che agiscono come ligandi oppure l’IL-6 che è importante per l’infiammazione.
Via JAK-STAT

Nel momento in cui si ha l’interazione con il ligando (IL-2, IL-3, IFNs, EPO, G-CFS, GH, prolattina), il recettore dimerizza ed è in grado di legare delle chinasi chiamate Jak (chinasi Janus associate). Quando Jak si avvicina, si auto-fosforila, diventa attiva e fosforila a sua volta il recettore a livello della porzione citoplasmatica. In questa porzione si ancora STAT (Signal Transducers and Activation of Transcription) che può essere fosforilata da Jak. La STAT fosforilata dimerizza e va a finire nel nucleo dove può regolare l’espressione dei geni che servono per la crescita cellulare.
Questa via è particolarmente importante per l’ematopoiesi, ovvero la produzione di globuli rossi. L’eritropoietina agisce con recettori di questo tipo.