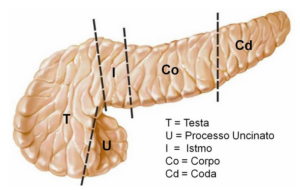
L’insulina, dal latino insula: isola, è uno degli ormoni più importanti del nostro organismo, prodotto e secreto dalle cellule β delle isole del Langerhans del pancreas.
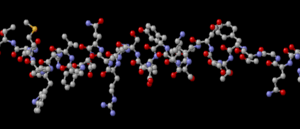
L’insulina è un polipeptide di 51 amminoacidi formato dall’unione di due catene, A e B di 21 e 30 amminoacidi, unite tra loro da 2 ponti disolfuro intercatena.
Uno fra la cisteina 7 della catena A e la cisteina 7 della catena B; e l’altro fra cisteina 20 della catena A e cisteina 19 della catena B.
Un ponte disolfuro intracatena presente nella catena A, fra cisteina 6 e cisteina 11.
I ponti disolfuro conferiscono all’insulina una struttura tridimensionale, da cui dipende la sua attività biologica.
L’insulina viene inizialmente sintetizzata (secondo meccanismi classi della sintesi proteica) in una forma detta “preproinsulina”, un polipeptide di 110 amminoacidi, in cui vi sono sequenze in più:
- Una sequenza “pre” aminoterminale (peptide-segnale, 24 aa), che consente la secrezione della proteina;
- Una sequenza centrale “pro” (peptide C, 35 aa), che determina il corretto ripiegamento del filamento polipeptidico.
Dopo la traduzione della preproinsulina nel reticolo endoplasmatico, un enzima taglia i 24 aa aminoterminali, lasciando la proinsulina, che si piega consentendo la formazione dei ponti disolfuro tra residui di cisteina. Quindi passa nelle vescicole dell’apparato del Golgi, dove il peptide C viene rimosso, formando così l’insulina.
L’insulina, viene immagazzinata, nella stessa cellula in forme di pacchetti polimerici, tenuti insieme da atomi di zinco, sotto forma di granuli secretivi che si accumulano nel citoplasma. (lo zinco è necessario per la stabilità strutturale della conservazione dell’insulina).
I granuli di insulina si accostano alla membrana cellulare, le membrane del granulo si fondono con la membrana cellulare liberando il pacchetto di insulina nell’interstizio;
l’ormone passa poi attraverso l’endotelio dei capillari entrando nel torrente sanguigno.
La secrezione avviene mediante esocitosi in risposta dei livelli di glicemia.
La cellula β presenta dei recettori specifici di membrana sensibili alle variazioni della glicemia.
Caricamento….
SECREZIONE INSULINICA
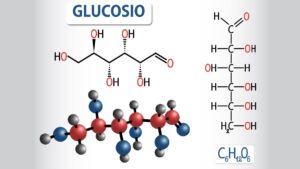
Una volta che il glucosio è in circolo, dopo l’assimilazione degli alimenti, tale molecola è alla base dei meccanismi che conducono al rilascio di insulina. Infatti si ritiene che il glucosio entri nella cellula beta ad opera di specifici gluco-trasportatori, in particolare GLUT2 e GLUT1; quindi il glucosio viene fosforilato dall’enzina glucochinasi e quindi avviato alla catena glicolitica.
Il piruvato che ne deriva entra nel mitocondrio attraverso il ciclo degli acidi tricarbossilici ed i successivi avvenimenti mitocondriali della catena respiratoria portano alla produzione di ATP.
L’aumento del rapporto ATP/ADP induce la chiusura a livello della membrana cellulare del canale del potassio ATP dipendente, cui consegue l’apertura dei canali del calcio voltaggio-dipendenti, con ingresso degli ioni calcio nella cellula.
Quest’ultimo meccanismo determina la liberazione di insulina attraverso la degranulazione.
Tale liberazione riconosce una prima fase rapida a cui segue una fase prolungata; inoltre la secrezione è pulsatile. Fino a quando la massa delle insule è conservata o, addirittura, accresciuta come nel giovane, tutto rientra nella fisiologia dei metabolismi. Quando, invece, si va avanti negli anni, allora i fenomeni apoptosici (morte programmata di una cellula) prevalgono sulla rigenerazione cellulare.

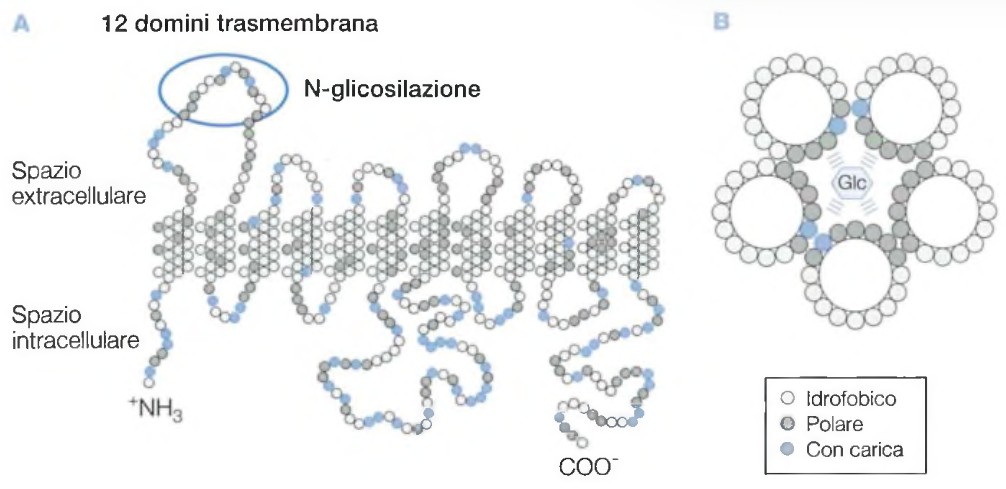
Tra tutti i gruppi di gluco-trasportatori, il più conosciuto e studiato è il GLUT4, molto sensibile all’insulina. Questo carrier, in condizioni normali, si trova in sede citoplasmatica e la sua traslocazione sulla membrana cellulare viene stimolata dal legame dell’insulina con il recettore di membrana.
Il recettore dell’insulina è una proteina chinasi specifica per i residui di tirosina. Il recettore ha 2 sub-unità (α e β). L’alfa sul lato extracellulare della membrana dove si lega l’insulina; il Beta attraversa la membrana.
Quando l’insulina si lega sul sito α, il sito β si autofosforila su residui di tirosina che a loro volta fosforilano le proteine bersaglio che sono dei secondi messaggeri. Questo processo favorisce lo spostamento del glucosio dal liquido interstiziale all’interno della cellula.
Quando la concentrazione ematica di glucosio si normalizza e l’insulina viene eliminata, le molecole di GLUT4 vengono lentamente rimosse dalla membrana plasmatica e sequestrate per endocitosi in vescicole intracellulari.
Il GLUT4 è presente soprattutto nel cuore, nel tessuto adiposo e nei muscoli scheletrici e vengono definiti insulino-dipendenti.

Normalmente il pancreas non produce insulina in maniera costante, bensì in quantità differenti, sostanzialmente in relazione con i pasti:
Now loading…
- Dopo un pasto ricco di carboidrati. La glicemia aumenta e il pancreas libera rapidamente insulina (picco insulinemico post-prandiale), che riporta la concentrazione del glucosio a livelli di normalità.
Il picco raggiunto è tanto più consistente quanto maggiori sono la quantità di zuccheri introdotta e la velocità del loro assorbimento (carico glicemico), mentre la presenza di fibra alimentare, rallentando l’assimilazione intestinale dei nutrienti, limita il picco di insulina.
Una volta soddisfatta la richiesta energetica della cellula dell’organismo, l’incremento di insulina stimola la conversione del glucosio in eccesso in depositi di glicogeno, in particolare a livello del fegato e dei muscoli. - Situazione normale di riposo (basale). Appena la glicemia comincia a diminuire, la secrezione di insulina si riduce rapidamente, tornando a valori simili a quelli del digiuno dopo 2 o 3 ore dall’assunzione del pasto.
- Breve periodo di digiuno. La produzione di insulina rimane a valori di base, senza picchi, nel digiuno notturno. A un certo punto, il calo della glicemia stimola le cellula α del pancreas a produrre glucagone, che provoca la conversione del glicogeno in glucosio e la liberazione di questo nel sangue. La richiesta minima di glucosio è pertanto assicurata dalle riserve di glicogeno. Il cervello riceve un segnale e lo traduce in sensazione di fame.
- Digiuno prolungato. Quando il digiuno perdura, le riserve di glicogeno dell’organismo si assottigliano gradualmente. La secrezione di glucagone è massima e aumentano altri ormoni (adrenalina, cortisolo) che innescano ulteriori meccanismi di approvvigionamento di glucosio. Esaurite le riserve di glicogeno, l’organismo attinge energia dai lipidi (attraverso la lipolisi) ed utilizza gli AA ottenuti dalla degradazione delle proteine per produrre glucosio (gluconeogenesi). Entrambi questi meccanismi sono a lungo andare deleteri per l’organismo, il primo (lipolisi) perché porta all’accumulo di corpi chetonici (chetoacidosi→coma→morte) e il secondo (gluconeogenesi) perché, una volta esaurite le proteine introdotte con gli alimenti, l’organismo arriva a degradare le proprie proteine strutturali (per esempio quelle muscolari).
- La lipolisi o catabolismo lipidico è un processo metabolico che prevede la scissione dei trigliceridi permettendo la liberazione degli acidi grassi liberi (FFA) e glicerolo (glicerina). La lipolisi è mediata dalla lipasi ed è indetta dagli ormoni glucagone, somatotropina (o GH), cortisolo, adrenalina, noradrenalina e testosterone.
- La gluconeogenesi è una via metabolica che permette di formare ex novo glucosio da molecole che non sono carboidrati. I precursori del glucosio sono soprattutto gli amminoacidi glucogenici (GAA) nel digiuno ed il lattato proveniente dai muscoli dopo un intenso sforzo muscolare e dagli eritrociti.
- La chetogenesi è una via sintetica che porta alla formazione dei corpi chetonici a partire dall’acetil-CoA. I corpi chetonici sono 3 composti che sono presenti nel sangue in piccole quantità. Questi composti sintetizzati dalle cellule epatiche in caso di eccesso di acetil-CoA sono: acetone, acido acetoacetico, acido beta-idrossibutirrico.
(PER NON FARSI IL SANGUE ACIDO)
- Sovralimentazione. In risposta all’eccessiva assunzione di cibo, soprattutto di alimenti ad alto contenuto di zuccheri, la produzione di insulina è intensamente e costantemente stimolata. L’organismo non ha modo di eliminare il glucosio, può solo utilizzarlo o accumularlo, quindi, una volta soddisfatta le richieste energetiche e saturate le riserve di glicogeno, il glucosio è convertito in un deposito energetico alternativo, ossia in acidi grassi, accumulati nel tessuto adiposo (grasso) sotto forma di trigliceridi. Questo processo conduce nel tempo a un incremento del peso corporeo.
- Sovralimentazione prolungata. Se si persiste nell’eccedere con il cibo, la produzione di insulina si mantiene a livelli costantemente elevati (iperinsulinismo), ma nel tempo le cellule dell’organismo perdono gradualmente la loro sensibilità all’insulina (insulino-resistenza) e ciò si traduce in un aumento permanente dei livelli di glucosio nel sangue.
Questa condizione si verifica nei soggetti obesi: anche se non tutti i soggetti obesi sono insulino-resistenti e non tutti quelli insulino-resistenti sono obesi, è accertato che tra obesità e alti livelli di insulina circolante esiste una stretta correlazione e che l’eccesso ponderale riduce la sensibilità dei tessuti periferici all’insulina.
Now loading…
La secrezione dell’insulina è regolata principalmente dalla glicemia, ma anche da altre sostanze come AA, ormoni (glucagone, cortisolo, somatotropine) o farmaci possono stimolarlo.
La maggior parte dell’insulina viene degradata nel fegato e nei reni grazie all’enzima glutadione insulina transidrogenasi e dall’enzima proteolitico tissutale specifico denominato insulinasi.
Le funzioni fisiologiche dell’insulina sono numerose; oltre alla riduzione della glicemia, l’insulina entra nel metabolismo proteico, nel metabolismo lipidico, nel metabolismo idro-salino, nell’attività cellulare e nella produzione di energia.
Gli organi bersaglio più importanti dell’insulina sono il fegato, il muscolo e il tessuto adiposo ma con importanti differenze funzionali:
- EFFETTI DELL’INSULINA SUL FEGATO: promuove la sintesi del glicogeno, incrementa la sintesi di trigliceridi e colesterolo, incrementa la sintesi proteica, inibisce la glicogenolisi, la gluconeogenesi e la chetogenesi.
- EFFETTI DELL’INSULINA SUL MUSCOLO: promuove la sintesi proteica, la sintesi del glicogeno, favorisce l’assorbimento di amminoacidi.
- EFFETTI DELL’INSULINA SUL TESSUTO ADIPOSO: promuove il deposito di trigliceridi, inibisce la lipolisi, favorisce l’assorbimento di glucosio ed acidi grassi.
Caricamento….