I tessuti epiteliali ghiandolari sono epiteli che svolgono una funzione di secrezione da parte di ghiandole che possono essere di due tipi: esocrine (come il fegato) che producono un secreto che viene riversato all’esterno o in ghiandole comunicanti con l’esterno dell’organismo; endocrine (come l’ipofisi) che producono un secreto che finisce nel sangue (gli ormoni).

Gli epiteli secernenti derivano da una invaginazione dello stesso epitelio di rivestimento dove poi riversano il loro secreto nel caso degli epiteli ghiandolari esocrini, per cui potrà avere derivazione ectodermica o endodermica. Il secreto viene riversato, attraverso il dotto escretore, in una zona esterna.
Le ghiandole esocrine mantengono un collegamento con l’epitelio di rivestimento di origine (il dotto escretore), mentre le ghiandole endocrine perdono il collegamento con l’epitelio di rivestimento di origine e riverseranno il secreto direttamente nel sangue.
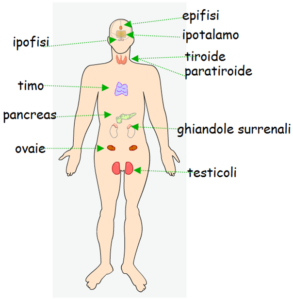
Esistono degli organi con funzione secernente formati da cellule connettivali che si trovano disposte nell’interstizio di organi come il testicolo (cellule di Leydig), dove producono l’ormone testosterone, e l’ovaio, dove producono ormoni come estrogeno e progesterone.
Esistono anche dei neuroni che svolgono la funzione secernente e producono altri ormoni come l’ossitocina, vasopressina, DOPA e serotonina, come quelli della neuro-ipofisi.
Now loading…
La ghiandola secernente, in genere, non è composta solo da cellule epiteliali, ma anche da altre cellule che servono di supporto alla funzione delle cellule secernenti, infatti gli epiteli, non essendo vascolarizzati, hanno bisogno di cellule connettivali che formano quello che si chiama stroma o tessuto interstiziale che garantiscono sostegno e nutrimento alla ghiandola. La parte invece epiteliale che produce il secreto prende il nome di parenchima.
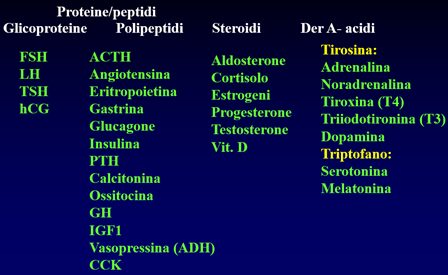
Il secreto prodotto dalle ghiandole può avere da un punto di vista chimico una natura diversa: muco-polisaccaridica (o zuccherina), peptidica, lipidica.
Oltre che per la zona di secrezione, le ghiandole endocrine e quelle esocrine differiscono tra di loro per la quantità del secreto prodotto. Le ghiandole endocrine producendo gli ormoni (messaggeri chimici) saranno di dimensioni più piccole ed il loro secreto sarà prodotto in piccole quantità.

Ghiandole esocrine
Gli epiteli ghiandolari esocrini sono classificati innanzitutto in unicellulari e pluricellulari e poi in base a 4 criteri:
- La posizione della ghiandola;
- La struttura istologica dei dotti escretori;
- La modalità di secrezione;
- Tipo di secrezione o tipo si secreto prodotto;
- In base alla posizione della ghiandola, le ghiandole pluricellulari extraparietali sono ghiandole esocrine di grandi dimensioni che fuoriescono dalla parete di un organo come il fegato e il pancreas, se sono incluse nella parete di un organo esso sono intraparietali. Queste si possono trovare o nello stesso tessuto epiteliale (intraepiteliali) oppure possono essere al di fuori del tessuto epiteliale (esoepiteliali) sotto l’epitelio di rivestimento. A loro volta le esoepiteliali possono essere coriali quando sono nel derma o nella tonaca propria, oppure sottomucose quando si trovano nell’ipoderma o nella sottomucosa.
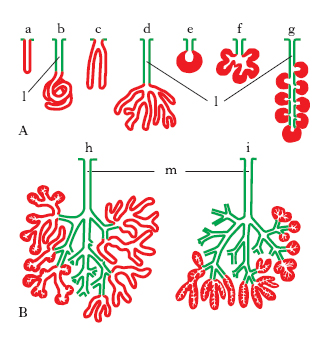
- In base alle caratteristiche del dotto escretore e delle cellule del parenchima si possono distinguere in semplici (A) e composte (B). Le semplici hanno un solo dotto escretore mentre le composte hanno un dotto escretore che si ramifica. A loro volta le semplici e le composte si dividono in altre categorie in base alla forma che assume la componente del parenchima che nella sua componente più piccola prende il nome di adenomero. L’adenomero è la più piccola parte della ghiandola, collegata ad un dotto escretore e costituita solo dalla parte secernente. Il dotto escretore che si collega al singolo adenomero si chiama dotto intercalare.
La forma dell’adenomero può essere a tubo (a) come le ghiandole che si trovano nell’intestino crasso del retto (Cripte del Lieberkühn), a tubo avvolto a gomitolo (b) come le sudoripare, a tubo che si ramifica (c – d), a forma d’acino o di alveolo (e) come la parotide e le ghiandole salivari, di acino o alveolo che si ramifica (f – g) come quelle nella trachea. Nelle composte c’è un’ulteriore forma che è quella mista fatta di tubi ed acini o alveoli (h). Se l’adenomero ha una forma sferica si chiama acino, se invece è sferico ma con una cavità molto ampia si chiama alveolo.
I dotti escretori sono alloggiati dentro il connettivo e paralleli ai dotti ci saranno i vasi sanguigni. Il tessuto connettivo avvolge la ghiandola creando una capsula. Da questo connettivo si dipartono dei setti connettivali che dividono la ghiandola in lobi e poi in strutture più piccole chiamate lobuli. Sia i lobi che i lobuli sono fatti dalla parte secernente (parenchima). In ogni lobulo di sono gli acini che costituiscono gli adenomeri. Nel tessuto connettivo di sostegno è contenuto un piccolo capillare sanguigno e poi il primo dotto escretore che viene dall’adenomero (dotto intercalare). Da questo dotto si arriva ad un dotto intralobulare ed infine al dotto escretore interlobare.
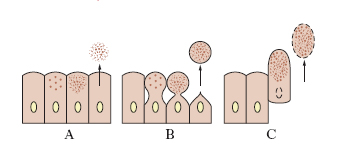
- In base al criterio di secrezione le ghiandole vengono suddivise in olocrine, apocrine e merocrine (ricordiamo che le cellule delle ghiandole esocrine sono fortemente polarizzate ed hanno una zona basale ed apicale completamente diversa perché la basale poggia sulla membrana basale, mentre quella apicale riversa il secreto verso il dotto escretore che è in comunicazione con l’esterno. Nelle ghiandole endocrine, invece, il secreto andrà verso il versante basale perché si riverserà nel sangue.)
La secrezione merocrina (A) consiste nel riversare il secreto in vescicolette che si fondono con la membrana apicale in modo simile all’esocitosi, per cui la cellula resta intatta. Nella secrezione apocrina (B) si assiste invece ad una specie di citodieresi, cioè si assiste alla divisione della cellula in due porzioni molto asimmetriche, in particolare l’estremità apicale si stacca dal resto. Per prima cosa si ha un accumulo di secreto nella porzione apicale, poi si ha la formazione di un restringimento ed alla fine si avrà il distacco della porzione apicale della cellula che andrà nel dotto escretore e poi all’esterno. Questa secrezione è tipica della ghiandola mammaria e della prostata. La secrezione olocrina (C) avviene in una modalità caratteristica in quanto la cellula si riempie di secreto, lo accumula sempre di più fino a quando non sarà completamente piena, il nucleo diventa picnotico e va in sofferenza, allora la cellula si stacca dalla membrana basale e, piena di secreto, va nel dotto escretore (la cellula stessa costituisce il secreto). Questa secrezione è tipica della ghiandola sebacea dove si ha un aspetto più acinoso. - In base al tipo di secreto prodotto possiamo individuare ghiandole mucose, sierose e miste. Il secreto mucoso è di natura glucidica (zucchero), il siero è invece di natura proteica, mentre il misto comprende entrambi. Alcune ghiandole tuttavia secernono anche lipidi, come la ghiandola sebacea.
Le ghiandole con secrezione proteica avranno per la maggioranza dei casi funzione enzimatica, le cellule che producono questo secreto saranno limitate da membrana sia per proteggere la cellula che per non disperdere le proteine idrosolubili nel citoplasma.

Ghiandole endocrine
La disposizione delle ghiandole endocrine è simile a quella delle ghiandole esocrine, vi è sempre una componente chiamata stroma costituita da tessuto connettivo e una parte secernente chiamata parenchima costituita da tessuto epiteliale. L’unica cosa che cambia è il luogo in cui viene riversato il secreto, ovvero nel sangue, ed avviene dal versante basale. Il criterio di classificazione delle ghiandole endocrine è più facile perché tiene conto della disposizione delle cellule secernenti all’interno della ghiandola stessa, perciò si dividono in tre categorie:
Caricamento…
- A cordoni o ammassi cellulari solidi, che comprendono quasi tutte le ghiandole endocrine (surrene, ipofisi, paratiroidi, pancreas endocrino).
- Ghiandole interstiziali, che hanno una struttura molto differente, caratterizzata da una dispersione di ghiandole endocrine all’interno di un organo che non è una ghiandola endocrina.
Queste ghiandole si trovano nel testicolo e nell’ovaio (nel loro interstizio) e sono le cellule del Leydig e le cellule accessorie del connettivo intorno al follicolo ovarico che producono rispettivamente testosterone ed estrogeni e progesterone. Anche le cellule parafollicolari della tiroide sono interstiziali. - Follicolari, in cui le cellule si dispongono a follicolo in maniera sferica e delimitano all’interno di questa sfera una cavità, come le cellule della tiroide (unico organo con questo aspetto).
Un’altra classificazione può essere fatta in base al secreto che viene prodotto, identificando due categorie:
- Ormoni proteici (messaggeri) che per la loro natura chimica non possono attraversare il doppio strato fosfolipidico della membrana della cellula bersaglio ed hanno quindi bisogno di un recettore specifico espresso sulla membrana basale della cellula.
- Ormoni steroidei (lipidici) che per la loro natura chimica passano tranquillamente la membrana della cellula bersaglio ed i ricettori sono presenti direttamente nel nucleo o nel citoplasma della cellula.
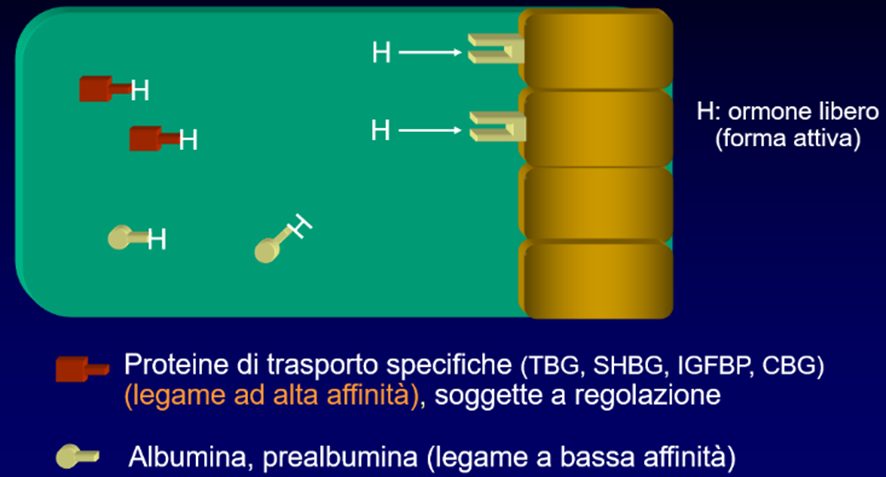
Il secreto della ghiandola endocrina, l’ormone, circola nel sangue perché deve raggiungere la cellula bersaglio ed è in genere legato a delle proteine carrier che ne facilitano il trasporto. Questo vale soprattutto per gli ormoni di natura lipidica, infatti, una volta legati alle proteine-vettore specifiche, sono trasportati fino alla cellula bersaglio dove si staccano dalla proteina carrier e attraversano la membrana della cellula bersaglio. Il recettore specifico si trova o nel citoplasma o nel nucleo. La reazione recettore-ormone determina un segnale nell’attività della cellula modificandone il comportamento: si altera l’espressione genica producendo una nuova proteina, o maggiore o minore produzione di una proteina.
Gli ormoni proteici, invece, non potendo attraversare la membrana, necessitano di un recettore specifico espresso sulla superficie della cellula bersaglio. A quel punto l’ormone produce un effetto sulla cellula bersaglio tramite la reazione recettore-ormone che innesca un segnale che porta una cascata fino al nucleo tramite secondi messaggeri come l’AMPc o Ca++ che attraverso delle chinasi portano all’attivazione di eventi post-trascrizionali o a espressione di geni cambiando la sintesi proteica della cellula (nella quantità o nella diversa produzione) nel nucleo.
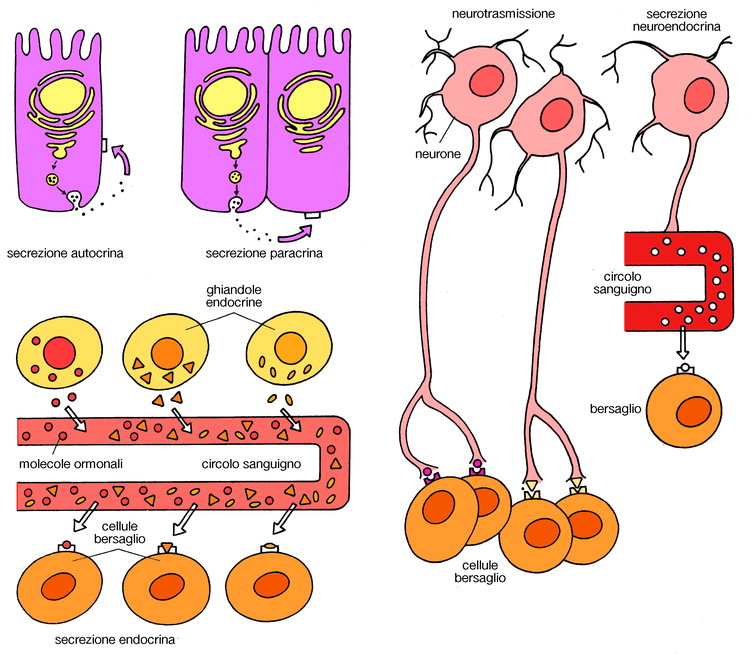
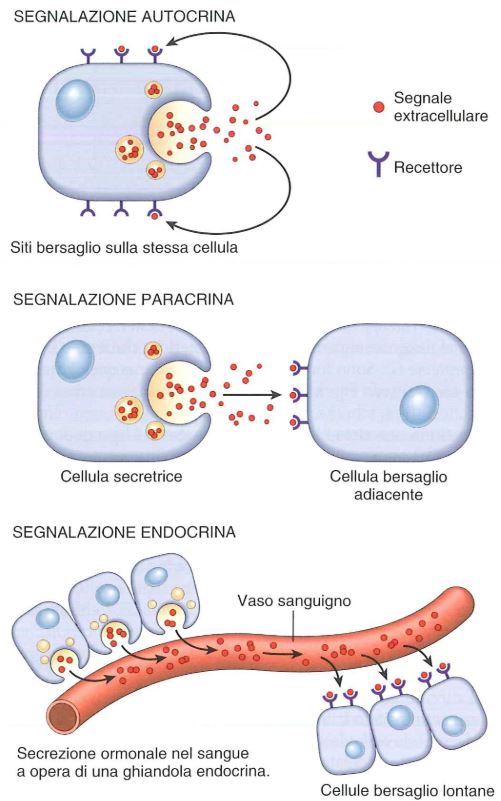
Il processo di secrezione viene definito come secrezione endocrina perché gli ormoni sono riversati nel sangue, però si possono avere anche processi di secrezione dove le cellule che producono l’ormone hanno come cellule bersaglio cellule molto vicine (secrezione paracrina), oppure processi di secrezione in cui la cellula che secerne è anche la cellula bersaglio (secrezione autocrina). Un altro tipo di secrezione è quella neuronale, operata da cellule nervose in cui l’ormone secreto è il neurotrasmettitore.
Un ormone su una cellula bersaglio può indurre anche una cellula a migrare, cambiando la conformazione del citoscheletro, tramite movimenti ameboidi.
Al microscopio elettronico una ghiandola endocrina a secrezione proteica è riconoscibile per vari aspetti: innanzitutto perché l’ormone è rivestito da una membrana, altrimenti il contenuto si disperderebbe nel citosol, inoltre sono cellule debolmente basofile perché gli ormoni sono prodotti in piccolissime quantità, il nucleo sarà di grandi dimensioni, così come i mitocondri e il RER. Anche l’apparato di Golgi, che adopera il processo di secrezione con una esocitosi sul versante basale, apparirà di grandi dimensioni.
Invece una ghiandola endocrina a secrezione lipidica non avrà un ormone rivestito da membrana, il citoplasma avrà un grande nucleo, un RER, mitocondri e lisosomi di grandi dimensioni. L’apparato di Golgi ed il REL sono coinvolti nella sintesi dei lipidi ed intorno alla cellula ci sono grosse quantità di colesterolo perché è la molecola che dovrà sintetizzare la cellula. La cellula, infatti, prende il colesterolo dal sangue, lo immagazzina nel suo citoplasma e poi con il REL ed i lisosomi lo trasforma per sintetizzare il granulo lipidico, cioè l’ormone steroideo. La sintesi che comincia nel REL si avvale dei lisosomi, continua nei mitocondri (per questo sono molto grandi, numerosi e con creste ben sviluppate) dove si avrà la maturazione dell’ormone steroideo.
Fonti: SlidePlayer – Treccani – Sapere