L’iperplasia è una forma di adattamento cellulare consistente nell’aumento del numero delle cellule. Si verifica come risposta adattativa in quegli organi in cui ci sono cellule che possono replicarsi. Con l’iperplasia aumenta il numero delle cellule e il volume dell’organo. Questi meccanismi sono armonicamente integrati in una risposta adattativa in cui, nel momento in cui cessa lo stimolo, tutto ritorna allo stato normale. Nelle neoplasie, che non sono risposte adattative, quei circuiti molecolari hanno vari punti in cui i segnali diventano costanti, come se il ligando fosse sempre attaccato al recettore.
L’iperplasia e l’ipertrofia possono coesistere, come nell’utero gravidico in cui vi è sia un’iperplasia, ovvero un aumento del numero di cellule dell’organo, sia una ipertrofia, un aumento delle dimensioni delle singole cellule (il lume intrauterino passa da un volume di 4-6 ml in una donna non gravida ad un volume di 4-5 litri in una donna gravida). Se consideriamo il miocardio, non abbiamo un’iperplasia, ma solo una ipertrofia, dovuta al fatto che le cellule non si possono replicare, quindi avviene solamente un aumento delle dimensioni della cellula stessa.
Iperplasia fisiologica
Dal punto di vista fisiologico si può avere un’iperplasia ormonale quando c’è un’alterazione della pressione ormonale durante la pubertà o la gravidanza. Si tratta di alterazioni del circuito ormonale che si traducono in una iperplasia della ghiandola mammaria, oppure dell’utero che durante la gravidanza diventa iperplasico ed ipertrofico. In questo caso l’iperplasia serve per aumentare le capacità funzionali del tessuto.
Rigenerazione epatica dopo epatectomia parziale. A. Lobi del fegato di ratto (M, lobo mediano; RL e LL, lobi laterali destro e sinistro; C, lobo caudato). L’epatectomia parziale rimuove due terzi del fegato (lobi mediano e laterale sinistro). Dopo 3 settimane il lobo laterale e il lobo caudato aumentano di volume fino a raggiungere una massa equivalente a quella del fegato originale, senza che si abbia però la ricrescita dei lobi mediano e laterale sinistro. B. Ingresso e progressione degli epatociti nel ciclo cellulare (A. Da Goss RJ: Regeneration versus repair. In Cohen IK et al [eds]: Wound healing. Biochemical e Clinical Aspects. Philadelphia, WB Saunders, 1992, pp 20-39). Fonte: Le basi patologiche delle malattie.
Un altro esempio classico di iperplasia fisiologica è l’iperplasia compensatoria, come quello che succede a livello del fegato. Infatti, in seguito ad asportazione di una porzione di fegato (epatectomia parziale), dopo un certo periodo, la porzione asportata sarà sostituita da nuovo tessuto epatico. Questo processo si chiama iperplasia compensatoria o iperplasia rigenerativa e serve per aumentare la massa tissutale di un organo dopo un danno o una parziale asportazione. Non avviene la ricrescita dei lobi asportati, ma avviene un aumento delle dimensioni dei lobi rimanenti (crescita compensatoria) che consentirà all’individuo di ripristinare la funzionalità del fegato anche se a livello morfologico la forma non sarà più quella iniziale. Le cellule sottoposte ad iperplasia rigenerativa sono principalmente gli epatociti che sono in fase G0, quindi sono delle cellule stabili che in questo contesto vengono stimolate ed entrano nella fase G1 del ciclo cellulare perché occorre rigenerare il fegato. Nei processi di iperplasia rigenerativa rientrano anche altre cellule, come le cellule stellate, anche se il meccanismo riguarda soprattutto gli epatociti.
Entrando nello specifico, durante l’iperplasia compensatoria si hanno delle alterazioni, ovvero un aumento delle proteine che sono coinvolte nella proliferazione cellulare, poi abbiamo la sintesi del DNA (che nell’epatocita normale non avviene) con un picco a 12-24 ore. Si ha proliferazione di epatociti, poi diminuzione della sintesi di DNA, e dopo 1-2 settimane la massa epatica è ripristinata. Quindi vi è una modificazione del comportamento delle cellule che da stabili entrano nel ciclo cellulare, vi è una modificazione del quadro dell’espressione genica degli epatociti e poi uno smorzamento del loro comportamento rigenerativo perché si è creato il tessuto epatico.
Durante le prime 4 ore, grazie all’intervento di mediatori di priming, si ha l’espressione di geni come c-fos e c-jun che servono per attivare la trascrizione dei geni della crescita cellulare e c-myc. A seguire, sotto l’influenza di altri mediatori, si ha l’espressione di altri geni come bclx, che ha una funzione anti-apoptotica, p53, mdm2 e poi delle cicline. Questo quadro ci dice che vi è un’armonica modificazione del quadro di espressione genica delle cellule affinché si abbia il processo di iperplasia rigenerativa.
Questa proliferazione dipende da fattori di crescita polipeptidici quali:
Fattore di crescita epatocitario (HGF) ed il suo recettore c-met, che è un recettore ad attività tirosin-chinasica intrinseca;
Fattore di crescita epidermico (EGF);
Fattore di crescita trasformanteα (TGF-α).
Durante il processo di rigenerazione epatica sono importanti anche delle citochine come l’interleuchina-6 (IL-6) ed il fattore di necrosi tumorale α (TNF-α). Mentre il TGF-α ha una attività autocrina sugli epatociti, gli altri mediatori agiscono in maniera paracrina. Le cellule non parenchimali del fegato, per esempio le cellule del Kupffer, liberano il TNF-α che è importante nel processo di rigenerazione.
La risposta genica durante la rigenerazione epatica prevede una risposta precoce immediata, in cui le cellule passano dallo stato di riposo G0 allo stadio G1. In questo stadio è importante l’attivazione di proto-oncogeni come c-fos e c-jun, che sono importanti per la trascrizione. C-fos e c-jun si uniscono (dimerizzano) e inducono il DNA alla trascrizione di geni che servono per la crescita cellulare. Questo perché agiscono sul fattore di trascrizione AP-1. Durante la risposta precoce immediata si ha anche l’intervento del proto-oncogene c-myc che viene espresso per la proliferazione cellulare. Esiste una seconda fase durante il progredire della rigenerazione epatica, che è la risposta genica precoce ritardata, in cui vi è l’intervento di bcl-x e mdm2 che impediscono l’apoptosi. Da un lato la risposta precoce immediata attiva la crescita cellulare, dall’altro attiva bcl-x che è anti-apoptotico.
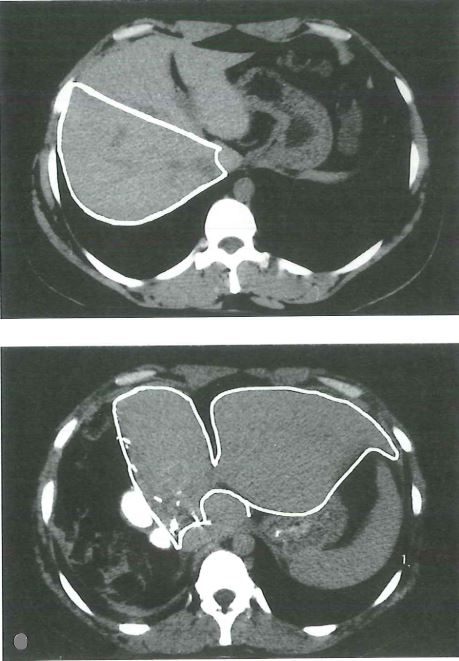
Rigenerazione del fegato umano nel trapianto da donatore vivente. Tomografia computerizzata (TC) del fegato del donatore dopo trapianto da donatore vivente. ll pannello superiore mostra una scansione del fegato del donatore prima dellintervento, evidenziando il lobo destro, che verrà usato per il trapianto. ll pannello inferiore mostra una scansione del fegato 1 settimana dopo l’epatectomia parziale. Da notare il notevole aumento di volume del lobo sinistro (evidenziato nel pannello), in assenza di ricrescita del lobo destro. (Per gentile concessione di: R. Troisi, MD, Ghent University, Ghent, Belgio). Fonte: Le basi patologiche delle malattie.
Nella fase di transizione da G1 a S, è importante il complesso ciclina D che si lega alla chinasi ciclina-dipendente 4 (CDK4). La ciclina D è importante per far progredire il ciclo cellulare legandosi al CDK4. Non solo succede questo, ma si ha la fosforilazione della proteina del retinoblastoma (Rb) da parte di CDK4 legata a ciclina D. In questo modo viene attivata la trascrizione di geni per la crescita cellulare, per esempio viene favorita la sintesi di ciclina E che interagisce con la CDK2.
Queste risposte a livello genico sono dovute a mediatori chimici importanti per far verificare il processo di rigenerazione epatica. Gli epatociti sono in G0 e devono entrare in G1, questo processo viene chiamato priming o innesco ed è favorito da mediatori come TNF-α e IL-6. Gli epatociti entrano nel ciclo cellulare e i fattori di crescita intervengono nella fase successiva che è quella di proliferazione regolata da fattori di crescita (HGF, TGF-α…) e da adiuvanti o co-mitogeni (norepinefrina, insulina, ormone tiroideo…).
Per quanto riguarda l’inibizione, oggi si sa che è importante il TGF-β. Il Transforming Growth Factor β è un gene oncosoppressore, il quale, quando secreto, è in grado di bloccare il ciclo cellulare interagendo con i recettori della superficie cellulare. In questo caso si ha l’inibizione del ciclo cellulare, l’abbassamento del tasso di crescita e una riduzione della domanda metabolica, quindi interviene nell’interruzione del processo di rigenerazione epatica.
Generalmente l’iperplasia rigenerativa inizia negli spazi periportali e poi si estende a livello di zone medio-lobulari e centro-lobulari. Se invece avviene un danno acuto, intenso da parte di un agente esterno che danneggia il parenchima epatico (come il tetracloruro di carbonio), la parziale iperplasia rigenerativa avviene solo a livello centro-lobulare, e non si riesce a rigenerare in maniera funzionale l’organo. Questa crescita parziale avviene grazie a delle cellule staminali specifiche dette cellule ovali. La stessa cosa avviene in caso di epatiti croniche.
Per quanto riguarda l’iperplasia patologica vi è per esempio quella dovuta ad alterata stimolazione ormonale, come l’iperplasia benigna della prostata, oppure l’iperplasia benigna dell’endometrio che è dovuta ad uno squilibrio tra il bilancio di estrogeni e progesterone, infatti si ha una prevalenza di estrogeni (che favoriscono la proliferazione cellulare) sul progesterone (che inibisce la crescita cellulare) che porta ad un’iperplasia dell’utero in quanto gli estrogeni iperstimolano l’endometrio e ciò è causa di sanguinamento endometriale. Le iperplasie, inoltre, possono essere indotte da alcune infezioni virali come il papilloma virus (HPV) che può indurre delle iperplasie a livello endocervicale provocando la formazione di verruche cutanee e lesioni mucose. Le formazioni verrucoidi sono i papillomi e devono essere controllate, se non direttamente asportate, perché rappresentano una condizione pre-neoplastica.









")




")
