Ogni componente cellulare ha la capacità di muoversi. I movimenti possono avvenire a livello subcellulare, quando riguardano gli organuli presenti all’interno della cellula (come i cromosomi in mitosi e il trasporto delle vescicole), a livello cellulare, quando riguardano gli spostamenti della cellula per intero (come nel caso dei protozoi ciliati, degli spermatozoi e delle metastasi), e al livello di tessuto e organismo pluricellulare, quando riguardano interi complessi cellulari (come accade per i muscoli scheletrici o il cuore). In genere l’energia impiegata per il movimento proviene dall’idrolisi dell’ATP o di qualche nucleoside trifosfato che determinano cambiamenti conformazionali di proteine motrici che, negli eucarioti, si associano a microtubuli e microfilamenti. Il movimento basato sui microtubuli riguarda interazioni tra microtubuli e le MAP motrici: dineine citoplasmatiche e assonemali e chinesine che consentono movimenti intracellulari e di appendici esterne. Il movimento basato sui microfilamenti riguarda interazioni tra microfilamenti di actina e le miosine come per quanto accade nelle fibre muscolari e nei movimenti non muscolari.
Motilità intracellulare
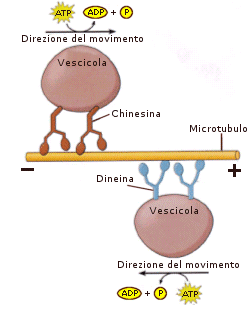
Le chinesine e le dineine si poggiano su un lato e l’altro del microtubulo e trasportano vescicole e strutture intercellulari sfruttando l’energia prodotta dall’idrolisi dell’ATP. Le chinesine trasportano le strutture verso le estremità positive dei microtubuli, cioè dal corpo cellulare verso la periferia, le dinesine si spostano invece nella direzione opposta, verso le estremità negative dei microtubuli, ovvero verso il centro della cellula dove è situato il centro di organizzazione dei microtubuli (MTCO).
Le chinesine sono organizzate in tre domini: una testa globulare (o motrice) che si lega al microtubulo e idrolizza l’ATP, una regione ad elica che rappresenta la parte centrale della chinesina ed uno stelo che ha la funzione di interagire con altre proteine ed organelli che costituiscono le molecole carico. Le teste globulari funzionano come piedi per “camminare” lungo il microtubulo verso l’estremità positiva. Per camminare, in seguito all’idrolisi dell’ATP, una testa motrice della molecola si distacca dalla molecola di tubulina e si sposta in avanti, seguirà il movimento della seconda testa che farà avanzare tutta la chinesina lungo il microtubulo.
Le dineine citoplasmatiche sono formate da associazioni di diverse catene: le catene pesanti che formano le teste globulari che interagiscono con il microtubulo, le catene intermedie e leggere che costituiscono la parte centrale della dineina e il complesso dinactina che consente di legare indirettamente la dineina alle membrane del cargo.
Le vescicole vengono trasportate dalle MAP motrici grazie alle vie costituite dai microtubuli che definiscono la polarità cellulare, la struttura a la distribuzione del reticolo endoplasmatico, dell’apparato di Golgi, dei lisosomi e delle vescicole di secrezione. Per quanto riguarda invece il trasporto assonale, le proteine motrici trasportano organelli e vescicole dal corpo cellulare verso estremità e viceversa sfruttando i microtubuli come piste.
Motilità cellulare
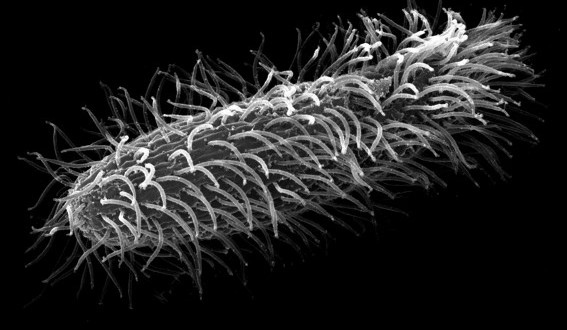
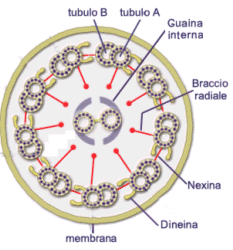
I microtubuli, però, consentono anche il movimento della cellula per mezzo di ciglia e flagelli attraverso il movimento di appendici mobili. Questi movimenti negli organismi unicellulari consentono lo spostamento da una parte e l’altra dell’ambiente e la raccolta del cibo come nel caso dei protozoi ciliati del tipo Paramecium. Negli organismi pluricellulari sono presenti solamente in cellule particolari e specializzate come gli spermatozoi che devono raggiungere la cellula uovo e le cellule dell’epitelio respiratorio per la raccolta del muco e della polvere. Nelle cellule, in genere, le ciglia sono più numerose, più corte ed hanno un movimento a “battito di remi”, mentre i flagelli sono più lunghi e radi ed hanno un movimento di tipo ondulatorio. La struttura portante di ciglia e flagelli è l’assonema, un sistema cilindrico di microtubuli rivestito di membrana plasmatica e connesso al corpo basale che funziona come MTCO. L’assonema è composto da varie coppie di microtubuli, tra le quali sono presenti delle proteine formate da dineina. Al centro di queste coppie vi è una decima coppia, per questo la struttura viene chiamata 9+2 (9 coppie periferiche e 2 microtubuli centrali). Alla base dell’assonema c’è il corpo basale con una struttura a raggiera di triplette di microtubuli in cui manca la coppia centrale e viene chiamata 9×3 (9 triplette di microtubuli in cerchio). I microtubuli dell’assonema vengono distinti in A e B. Da ciascun tubulo A delle doppiette esterne partono 2 bracci laterali (interno ed esterno) di dineina assonemale, che contatta il tubulo B della doppietta accanto e bracci radiali che collegano le doppiette alla coppia centrale. Ogni braccio è disposto in intervalli regolari nel senso longitudinali. La forza meccanica per scorrimento dei microtubuli data dall’idrolisi dell’ATP e cambiamenti conformazionali della dineina che fa scivolare due doppiette periferiche una sull’altra porta ad una flessione locale dell’assonema che poi genera il classico movimento ciliare o flagellare.
Altri tipi di movimenti importanti a livello di tessuto sono quelli basati sull’interazione tra actine e miosine. Le miosine sono coinvolte in processi diversi come la contrazione muscolare, il moto cellulare, la fagocitosi, le strutture uditive, il trasporto di melanina, ecc. Le miosine sono organizzate in una catena pesante che forma la testa globulare che lega actina e idrolizza l’ATP, e una zona fibrosa, chiamata coda, variabile che interagisce con altre molecole. Alcune miosine, come la miosina I, hanno una sola testa, altre, come la miosina II e la miosina V, si associano attraverso le code in un dimero a due teste. Le miosine di tipo II, sono quelle più diffuse e si trovano nel muscolo liscio e scheletrico, nel miocardio e in alcune cellule non muscolari.
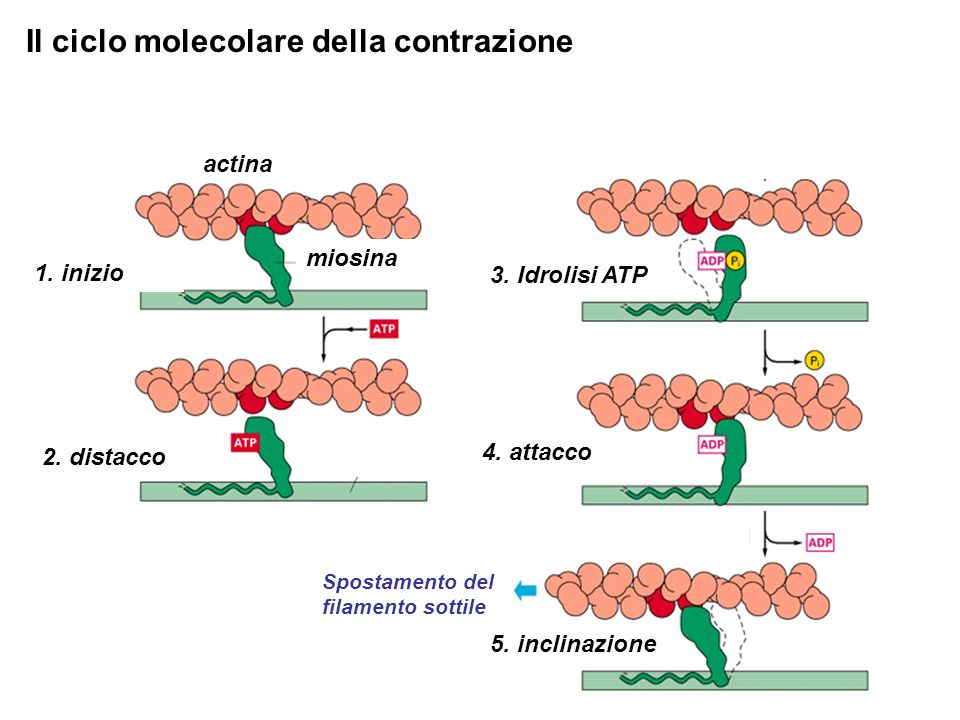
Actina e miosina sono disposte in maniera ordinata nei sarcomeri, le unità funzionali (contrattili) delle miofibrille, o fibre muscolari. La miofibrilla consiste di filamenti spessi e sottili intercalati. I filamenti sottili sono disposti ad esagono attorno ai filamenti spessi. I filamenti spessi delle miofibrille consistono di centinaia di molecole di miosina disposte in una struttura sfalsata ripetitiva. Le singole molecole di miosina sono inserite nel filamento longitudinalmente, con le loro teste globulari che sporgono dal centro del filamento. La porzione centrale del filamento contiene una zona centrale nuda priva di teste.
La contrazione è spiegata dal modello dello scorrimento dei filamenti: i filamenti spessi scorrono su quelli sottili trainandoli verso il centro del sarcomero che si restringe insieme alla banda I. Quindi i filamenti non si accorciano, ma aumenta la loro sovrapposizione! La contrazione muscolare è regolata dalla disponibilità di Ca2+ che è immagazzinata nel reticolo endoplasmatico, o meglio, il reticolo sarcoplasmatico, e il cui rilascio si ha in seguito ad un impulso nervoso. In assenza di ioni calcio, la testa della miosina non riesce ad attaccarsi sulla miosina. Quando invece il calcio si lega alla tropomiosina C, la miosina può legarsi all’actina.
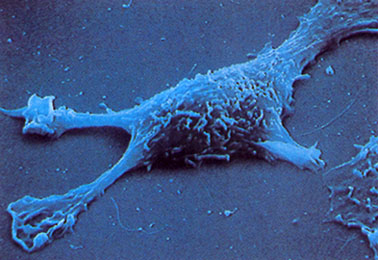
L’interazione actina-miosina è coinvolta anche in molti tipi di movimento non muscolare come la citocinesi, il movimento cellulare strisciante (ameboide e filopodi), la chemiotassi e le correnti citoplasmatiche. Il movimento strisciante è dovuto all’emissione di speciali prolungamenti citoplasmatici lamellipodi e filipodi. Il movimento mediato da filopodi è tipico dei fibroblasti e delle migrazioni cellulari embrionali. I filopodi emergono evidenti dalla superficie cellulare e al loro interno si trovano i fasci paralleli di actina. Così, le teste miosiniche si associano a questi microfilamenti di actina che man mano si allungano all’estremita positiva per polimerizzazione. I microfilamenti formano anche delle interazioni stabili che permettono un’adesione stabile al substrato tramite proteine integrine. Il movimento strisciante di tipo ameboide consiste nelle emissioni di protrusioni citoplasmatiche dette pseudopodi e nell’innestamento di una corrente citoplasmatica tramite l’alternanza di transizioni gel-fluido. Il citoplasma tende a diventare più fluido verso il pseudopodio ed a solidificarsi sulla punta mentre, al margine posteriore, l’ectoplasma fluidifica e scorre verso la protrusione. Questo tipo di movimento è tipico di alcuni protozoi, funghi e leucociti e viene utilizzato anche per la fagocitosi.