Gli epiteli pluristratificati si definiscono in base all’ultimo strato di cellule sul versante apicale.
L’epitelio pluristratificato non corneificato (o non cheratinizzato) è chiamato così perché le cellule fino all’ultimo strato hanno il nucleo. Questo epitelio è presente nel cavo orale, nella cornea, nell’esofago e nella vagina. Sono tonache mucose, in particolare quella della vagina presenta cellule molto grandi con un citoplasma molto trasparente. Se si passa dallo strato basale a quello apicale, c’è un cambiamento di forma delle cellule: da più piccole diventano grandi, si differenziano e solo negli ultimi strati diventano appiattite (pavimentose). Tutte le cellule degli strati centrali hanno un citoplasma molto chiaro e le cellule sono di grandissima dimensione. Queste tonache mucose differiscono dall’epidermide per la presenza del muco.

L’epidermide, però, è diverso dagli epiteli pavimentosi stratificati non cheratinizzati perché le cellule degli ultimi strati non si riconoscono, ma si ammassano le une sulle altre, i nuclei non sono più visibili ed è definito epitelio pavimentoso composto cheratinizzato perché le cellule esterne sono fatte quasi totalmente di cheratina (proteina principale contenuta nelle cellule dell’epidermide) e corneificato perché c’è uno strato di cellule ammassati che hanno perso i nuclei e non hanno una netta distinzione tra le membrane cellulari perché si sono fuse. Questo strato corneo può avere spessore variabile a seconda del distretto anatomico interessato (ad esempio nei palmi delle mani e dei piedi è più spesso) e in base alla resistenza meccanica che è necessaria. Le cellule apicali sono prive di nucleo perché sono cellule morte in quanto sono lontane dal connettivo, fonte di nutrimento. Le cellule basali, invece, sono cubiche, molto vicine tra di loro e vanno incontro a mitosi.
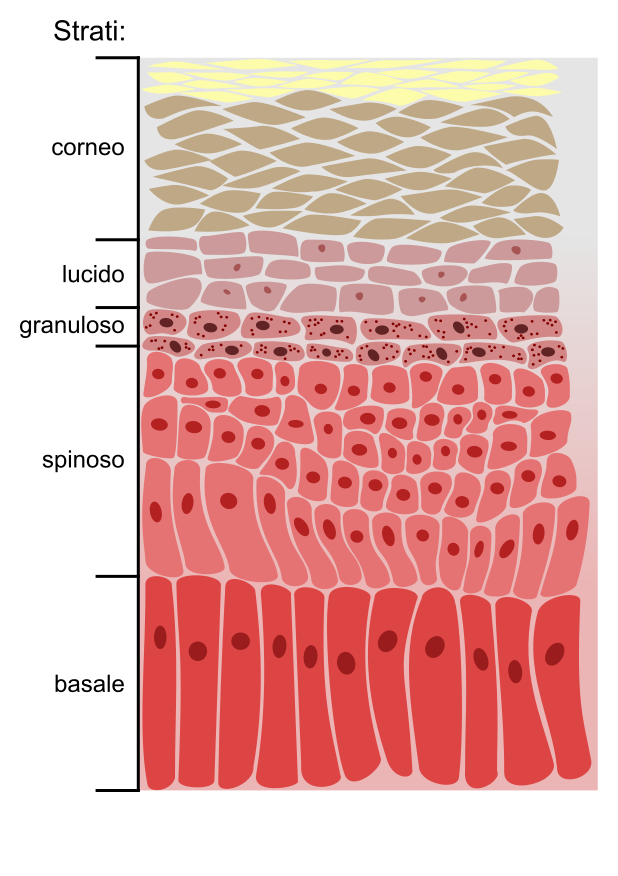
Lo strato basale, in realtà, è un insieme di 3-5 strati di cellule che poggiano sulla membrana basale con gli emidesmosomi (giunzioni ancoranti) ed integrine. Al di sopra dello strato basale si hanno cellule differenziate formate dagli strati spinosi o di Malpighi (4-8 strati) dove le cellule iniziano a cambiare forma emettendo dei prolungamenti, i tonofilamenti, coinvolti nelle giunzioni cellulari, che poi costituiscono delle fibrille (tonofibrille) che si impacchettano l’una sull’altra a ridosso del versante esterno della membrana plasmatica ed iniziano ad accumulare cheratina con cui entrano in contatto con le altre e cominciando ad accumulare cheratina (per questo le cellule si chiamano cheratinociti) fino ad assumere le caratteristiche finali delle cellule degli strati apicali. Il deposito di cheratina sul versante interno della membrana plasmatica è aiutato dalla proteina involucrina che lega le tonofibrille l’una all’altra per condensare la cheratina intorno alla membrana plasmatica del cheratinocita. Inoltre, all’interno dei cheratinociti dello strato spinoso, sono visibili dei granuli di due tipi: i melanosomi ed i cheratinosomi. I cheratinosomi sono fatti di lipidi che vengono secreti dai cheratinociti negli spazi intercellulari per condensare le cellule tra di loro ed impermeabilizzare l’involucro cellulare finale.

Dopo lo strato spinoso c’è lo strato granuloso, in cui all’interno delle cellule sono presenti dei granuli di una sostanza chiamata cheratoialina costituita da filagrina e loricrina che servono a formare le tonofibrille fatte di cheratina che rendono le cellule più schiacciate.
Nelle zone in cui l’epidermide è più spesso (palmo della mano e pianta del piede) esiste anche lo strato lucido, caratterizzato dal fatto che le cellule cominciano a perdere i nuclei e si colorano molto meno, questo perché esse da basofile diventano acidofile in quanto nelle cellule che sintetizzano cheratina sono abbondanti i ribosomi (fatti da acido ribonucleico).
L’ultimo strato (strato corneo) è formato da cellule che si fondono tra di loro diventando un ammasso cellulare pieno di cheratina con funzione di protezione e ricche di giunzioni intercellulari del tipo dei desmosomi. L’integrità dell’ammasso è garantita anche dalla presenza dei cheratinosomi negli spazi intercellulari. È questo ammasso proteico-lipidico che garantisce resistenza meccanica e impermeabilizzazione all’epidermide. L’impermeabilizzazione è dovuta soprattutto alla presenza dell’idrossicerammide, una sostanza lipidica di natura sierosa.
Le cellule che costituiscono l’epidermide sono prima di tutto i cheratinociti che sono tenuti insieme da giunzioni cellulari del tipo dei desmosomi (emidesmosomi). Quest’ultimi sono delle strutture che utilizzano anche proteine di membrana e glicoproteine di membrana che sono delle integrine della famiglia della α6β4. Le integrine fanno parte della membrana plasmatica ed hanno un dominio extracellulare che serve a determinare l’interazione della cellula con il substrato, quindi rappresentano un’importanza strategica nel collegamento con i cheratinociti dello strato basale e la membrana basale nel caso dell’epidermide. Il fatto che l’integrina sia α-β vuol dire che essa è un eterodimero costituito da due catene diverse. La mancata o trasformata espressione di queste integrine crea dei problemi gravi nel processo di adesione dei cheratinociti alla membrana basale o nei processi di differenziamento del cheratinocita, come nel caso delle neoplasie.

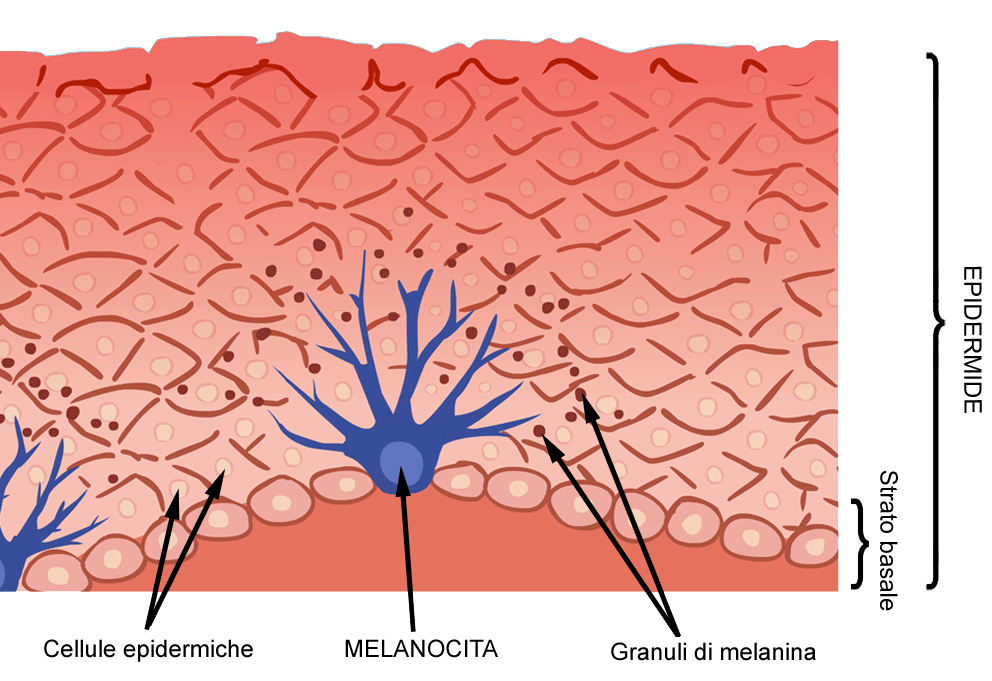
Oltre ai cheratinociti, nell’epidermide ci sono anche i melanociti che derivano da una porzione particolare del mesenchima ovvero le creste neurali (neuroectoderma), i cheratinociti derivano invece dall’ectoderma. I melanociti sono cellule molto grandi e localizzati prevalentemente a livello dello strato basale e nei primi strati dello strato spinoso. Essi emettono dei prolungamenti cellulari molto lunghi (dendriti) che si aggrappano a circa 30 cellule costituendo l’unità melanica, in particolare le porzioni terminali di questi prolungamenti si staccano, costituiscono delle vescicole di derivazione cellulare (i melanosomi). I melanosomi iniziano a vedersi nei cheratinociti dello strato spinoso fino agli strati successivi, hanno una struttura lamellare concentrica e sono corpuscoli all’interno del quale vi è la melanina. Quindi la melanina viene prodotta dal melanocita, si condensa in corpuscoli cellulari che si staccano dal melanocita stesso e vengono endocitati nei cheratinociti dello strato spinoso. Ovviamente questo è un fenomeno che avviene in dipendenza di stimoli dell’ambiente esterno (in risposta di esposizione a radiazioni ultraviolette). L’esposizione a raggi UV comporta il rilascio di MSH (ormone melanocita stimolante) che stimola la produzione di melanosomi che dà inizio al processo di citocromia tramite l’attivazione all’interno del melanosoma di un enzima che si chiama tirosinasi che trasforma la DOPA (diidrossifenilalanina) in melanina (fenomeno dell’abbronzatura). Esistono due forme di melanina, l’eumelanina che è un pigmento marrone-nero prodotto da coloro che hanno una colorazione più scura (fotoprotettiva in quanto garantisce protezione da raggi UV) e la feomelanina, prodotta dai soggetti più chiari con una colorazione giallo-rossa e che è dannosa (fototossica) e non protettiva.
L’esposizione ai raggi UV può generare neoplasie, perciò il nostro organismo produce uno schermo di protezione rappresentato dalla melanina che garantisce l’abbronzatura.
L’abbronzatura è una colorazione della pelle che può essere duratura (esposizione prolungata) con colorazione marrone o temporanea che provoca una colorazione della pelle arrossata che dipende dall’aumento del flusso sanguigno nelle cellule e non dai melanosomi che invece rimangono localizzati solo nell’estremità dei melanociti senza finire all’interno dei cheratinociti.
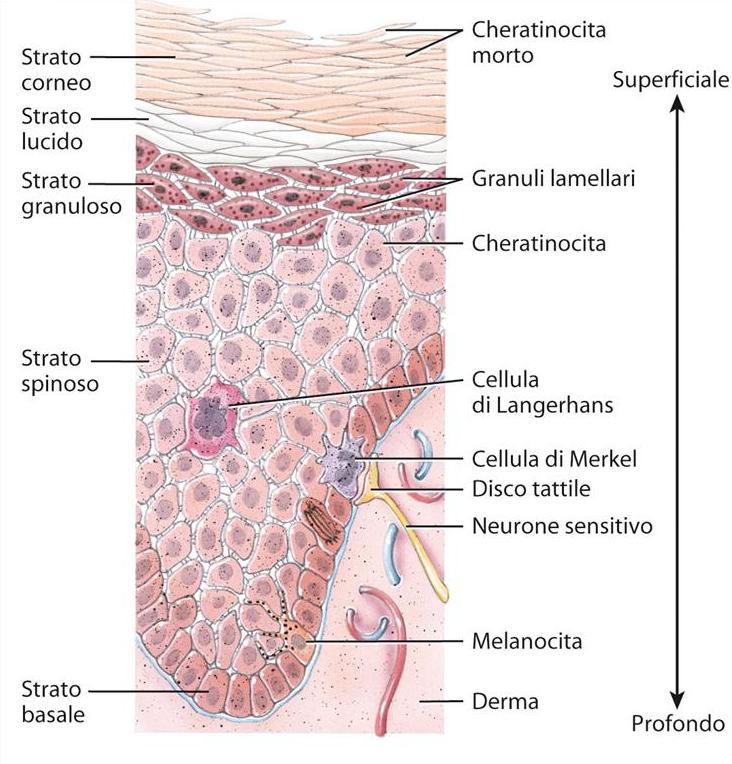
Le cellule di Langherans sono altri tipi di cellule presenti nell’epidermide che occupano lo strato spinoso, hanno una derivazione mesodermica, presentano dei prolungamenti ed appartengono alla linea monocita-macrofagica per cui svolgono una funzione di difesa, la prima forma di difesa. Esse non hanno delle funzioni fagocitarie dirette come i macrofagi standard, però guidano la difesa del nostro organismo producendo una sostanza attiva nel guidare la risposta di difesa, la citochina interleuchina 1 (IL-1). In particolare la IL-1 attiva la chemiotassi nel tessuto di cellule che vengono dal sangue, cioè di altri globuli bianchi, attirati ed attivati dalla secrezione di IL-1 da parte delle cellule di Langherans. Queste cellule sono mobili e perciò quando inglobano un antigene o microrganismo che penetra nella cute, si spostano nei linfonodi per presentare l’antigene ai linfociti B ed istruirli a distruggere quel determinato microrganismo, per le cellule di Langherans vengono classificate come APC (Antigen Presenting Cells).
Le cellule di Merkel derivano dal neuroectoderma, si trovano nello strato basale dell’epitelio e contengono la precheratina che permette di formare dei desmosomi e riesce a legarsi a tutte le cellule circostanti (è molto simile alle cellule dello strato basale) e di scambiare messaggi. Infatti possiede i Granuli di VIP e metaencefalina che agiscono da neurotrasmettitori. In seguito ad uno stimolo tattile, grazie al sistema dei desmosomi, il messaggio arriva alla cellula di Merkel che rilascia VIP e metaencefalina che trasmettono il messaggio alle terminazioni nervose sottostanti che fanno arrivare lo stimolo al cervello.
Il cavo orale è particolare perché è paracheratinizzato, in quanto pur avendo un accumulo di cheratina, le cellule degli ultimi strati non muoiono e mantengono i nuclei.
Fonti: Wikipedia – Scuola Anatomia – Pura Pelle